Физика/Применение физических методов медицине
Ушенко
Є.Г., Ушенко Ю.О.
Буковинський державний медичний
університет
Поляризаційні сингулярності
лазерних об’єктних полів біологічних тканин
1. Умови формування поляризаційних сингулярностей у лазерних
зображеннях біологічних тканин
Відомо, що
оптичні властивості біологічних тканин можна змоделювати сукупністю одноосних
двопроменезаломлюючих кристалів, що характеризуються матрицею Джонса.
Елементи
матриці когерентності 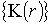 в кожній точці
в кожній точці  зображення
біологічної тканини можуть бути взаємозв’язані з її оптичною (координатні
розподіли фазових зсувів
зображення
біологічної тканини можуть бути взаємозв’язані з її оптичною (координатні
розподіли фазових зсувів  ) та геометричною (координатні розподіли напрямків оптичних
осей двопроменезаломлюючих фібрил
) та геометричною (координатні розподіли напрямків оптичних
осей двопроменезаломлюючих фібрил 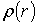 ) структурами біологічних тканин. На основі визначеного таким
чином взаємозв’язку відкривається можливість виявлення основних умов формування
лінійних (
) структурами біологічних тканин. На основі визначеного таким
чином взаємозв’язку відкривається можливість виявлення основних умов формування
лінійних (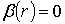 ) та циркулярних (
) та циркулярних (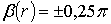 ) станів поляризації в різних точках зображення біологічної
тканини. Інакше кажучи, процес взаємодії лазерного випромінювання з сукупністю
оптично одноосних анізотропних фібрил можна розглядати як процес генерування
поляризаційних сингулярностей двопроменезаломлюючим біологічним об’єктом.
) станів поляризації в різних точках зображення біологічної
тканини. Інакше кажучи, процес взаємодії лазерного випромінювання з сукупністю
оптично одноосних анізотропних фібрил можна розглядати як процес генерування
поляризаційних сингулярностей двопроменезаломлюючим біологічним об’єктом.
Визначимо
умови формування сингулярних станів поляризації у зображенні такого об’єкту:
1) умови
формування лінійних ( ) поляризаційних сингулярностей
) поляризаційних сингулярностей
 (1)
(1)
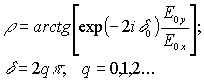 (2)
(2)
2) умови
формування циркулярних (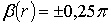 ) поляризаційних сингулярностей
) поляризаційних сингулярностей
 , (3)
, (3)
 . (4)
. (4)
Експериментально
координатні розподіли таких точок можна встановити шляхом виборки з усіх
можливих станів поляризації (поляризаційне картографування біологічних тканин)
розподілів лінійних і циркулярних станів поляризації в різних точках зображення
біологічної тканини.
2. Поляризаційно-сингулярна диференціація
біологічних тканин
В якості
об’єктів дослідження використовувалися оптично тонкі (коефіцієнт ослаблення 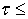 0.1) гістологічні
зрізи здорової (Рис. 1а,б) і патологічно зміненої (рання стадія колагеноза –
розростання сполучної тканини) нирки (Рис. 1в,г).
0.1) гістологічні
зрізи здорової (Рис. 1а,б) і патологічно зміненої (рання стадія колагеноза –
розростання сполучної тканини) нирки (Рис. 1в,г).

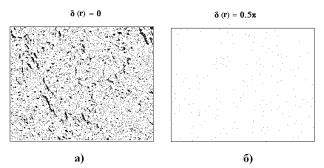
Рис. 1 Поляризаційні зображення гістологічних зрізів
фізіологічно нормальної (a, б) і патологічно зміненої (в, г) тканини нирки.
Фрагменти (a, в) відповідають ситуації співвісних поляризатора й аналізатора;
(б, г) – ситуації схрещених поляризатора й аналізатора
Проведемо
порівняльний статистичний аналіз координатних розподілів сингулярних точок
даних зображень (Рис. 2 і Рис. 3).

Для аналізу
даних розподілів на Рис 4 показані кількості 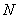 сингулярно
поляризованих точок зображень фізіологічно нормальної і патологічно зміненої
тканини нирки відповідно.
сингулярно
поляризованих точок зображень фізіологічно нормальної і патологічно зміненої
тканини нирки відповідно.
Рис. 5.11 Кількості  сингулярно
поляризованих точок (
сингулярно
поляризованих точок (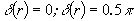 ) зображення фізіологічно нормальної (а) та патологічно
зміненої (б) тканини нирки.
) зображення фізіологічно нормальної (а) та патологічно
зміненої (б) тканини нирки.
З отриманих
даних видно, що залежності 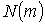 , визначені для зображень зразків тканини нирки обох типів,
мають складний, випадковий характер.
, визначені для зображень зразків тканини нирки обох типів,
мають складний, випадковий характер.
Більш
детальну кількісну інформацію про характер розподілів 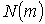 зображень тканини
нирки з різним фізіологічним станом несуть статистичні моменти першого
зображень тканини
нирки з різним фізіологічним станом несуть статистичні моменти першого  , другого
, другого 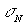 , третього
, третього  й четвертого
й четвертого 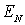 порядків (Табл. 1).
порядків (Табл. 1).
Таблиця 1 Статистичні моменти 1-4 порядків
розподілу поляризаційних сингулярностей зображень тканини нирки
|
Зразок тканини
|
Норма
(37 зразків)
|
Патологія
(36 зразків)
|
|

|
0.634 5% 5%
|
0.706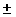 7% 7%
|
|
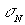
|
0.198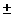 4% 4%
|
0.149 6% 6%
|
|

|
2.689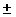 12% 12%
|
21.75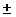 15% 15%
|
|

|
3.8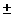 14% 14%
|
46.8 18% 18%
|
Видно, що
статистичний розподіл сингулярно поляризованих точок у зображенні здорової
тканини нирки близький до нормального - величини асиметрії 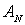 й ексцесу
й ексцесу 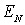 розподілів
розподілів  досить малі, а для
зображенні патологічно зміненої тканини нирки ці значення зростають практично
на 1 порядок.
досить малі, а для
зображенні патологічно зміненої тканини нирки ці значення зростають практично
на 1 порядок.
З іншого боку, з робіт Мандельброта відомо, що біологічні
об’єкти мають самоподібну структуру. Тому актуальним виявилося дослідження
самоподібності біологічних тканин при зміні їх архітектонічної складової.
Рис. 12 ілюструє залежності 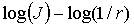 , отримані для координатних розподілів кількості
, отримані для координатних розподілів кількості 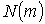 поляризаційних
сингулярностей зображень патологічно зміненої (а) і фізіологічно нормальної (б)
тканини нирки.
поляризаційних
сингулярностей зображень патологічно зміненої (а) і фізіологічно нормальної (б)
тканини нирки.

Рис. 12 Log-log залежність спектра потужності  розподілу кількості
розподілу кількості 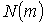 поляризаційних
сингулярностей зображення патологічно зміненої (а) і фізіологічно нормальної
(б) тканини нирки.
поляризаційних
сингулярностей зображення патологічно зміненої (а) і фізіологічно нормальної
(б) тканини нирки.
Апроксимація даних залежностей 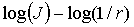 методом найменших
квадратів для поляризаційних сингулярностей зображень патологічно зміненої
тканини нирки дає набір ламаних ліній з багатьма значеннями локальних нахилів
методом найменших
квадратів для поляризаційних сингулярностей зображень патологічно зміненої
тканини нирки дає набір ламаних ліній з багатьма значеннями локальних нахилів  (Рис.12 а).
(Рис.12 а).
Поляризаційно сингулярна структура зображення
фізіологічно нормальної тканини самоподібна (фрактальна) – статистична обробка
залежностей 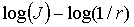 дає пряму лінію з
одним нахилом (Рис. 12 б).
дає пряму лінію з
одним нахилом (Рис. 12 б).
Отже, визначено аналітичні умови формування лінійних і
циркулярних поляризаційних сингулярностей і проаналізовано експериментально
одержані статистичні характеристики їхніх координатних розподілів у зображеннях
біологічних тканин різної морфологічної будови і фізіологічного стану.
Показано, що найбільш чутливими до зміни
оптико-геометричної структури біологічної тканини є 3-й і 4-й статистичні
моменти розподілів 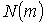 .
.
Встановлено, що координатна структура поляризаційних
сингулярностей зображення патологічно зміненої біологічної тканини є
випадковою, а фізіологічно нормальної – фрактальною.