Биологические
науки /9. Биохимия и биофизика
Candidate
of Biology E. V. Pinyaskina
Establishment of the
Russian Academy of Sciences Pricaspian institute
of Biological resources
of the Dagestan centre of science of the RAS
Protective
photoeffects induced by low-intensive red light
At present, the known types of photoinduced
cell reactivation and defense [5,8] are aimed at the removal or prevention of
one type of lethal photoproducts, namely, pyrimidine dimers, which are formed
in DNA affected by far- and middle- wavelength ultraviolet (UV) radiation
(220-320 nm). So far, no data is available on the photoinduced increase in
viability of cells, inactivated by near-UV radiation (320-380 nm) and visible
light (400-600 nm). Contrary to short wavelength UV radiation, the lethal
effect of optical radiation of these types occurs via a photodynamic mechanism
involving endogenous sensitizers, which generate reactive oxygen species that
induce DNA damage other than that of pyrimidine dimers [1,6,7]
The purpose of this study was to reveal the
effects of photorecovery in yeast cells during their photodynamic inactivation
by optical radiation at wavelengths of 320380 nm and 400-600 nm.
Our studies of the effect of monochromatic
light at 400-730 nm on C. guilliermondii cells, which were
inactivated by near UV radiation (320-380 nm), have shown that the light of the
red part of the spectrum (610, 630, 660, 680, 710 nm) significantly increases
their viability. As has been established (data not shown), the maximal
activity in the expression of yeast photoreactivity is exerted by light at 680
nm; we shall designate it further as FR680.
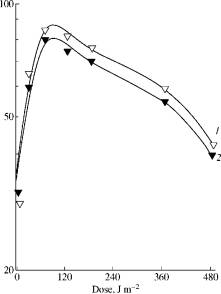
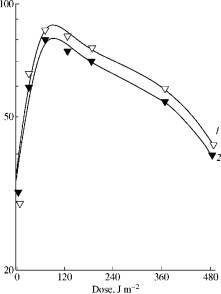
The typical curve characterizing the effect of red light (680 nm) on the
photoreduction of cell viability is shown at Fig.1. The maximal level FR680 is
reached already at short-term (several minutes) irradiation by monochromatic
light at low dose. The increase in time radiation.
|
Fig. 1. The effect of
monochromatic (680 nm) light irradiation dose at 22°C (1) and at 4°C (2) on
photoreduction of C. guilliermondii cells inactivated
by near UV (320-380 nm; 70 kJ/m2). |
|
Viability, %
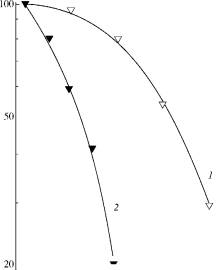
0 25 50 75 Dose, kJ m 2 Fig. 2. Viability curves
for S. cerevisiae irradiated by near UV (320-380 nm): (1)
wild type strain; (2) mutant strain rad 50-1. |
It is necessary to mention that the established shape of the FR680
dose curve mirrors the shape of corresponding curves that we obtained in our
studies on the resuscitation induced by monochromatic red light in yeasts C.
guilliermondii, previously inactivated by middle wavelength UV
radiation (290-320 nm) [4]. In addition we have shown that the efficiency of FR680
in the near UV for cell inactivation is not affected by the temperature
decrease to 4°C during monochromatic irradiation (Fig. 1). The same was also
observed previously in case of middle wavelength UV radiation [4].
These data suggest that in the
photorecovery of cells from the lethal effect of near UV radiation, the same
system is responsible - sensitive to red
light and involved in the effect of middle wavelength radiation. This further indicates the efficiency
of this system in respect to both pyrimidine dimers and photodynamic DNA
damage, such as monostrand breaks, which
under near UV radiation are important for cell inactivation [1,7].
It is well established that UV-induced damage
can be repaired mainly by excision and post-replicative DNA reparation systems.
Therefore it was worth to examine the possible association between FR680
radiation and photoinduced activation of these reparation systems.
|
Dose, kJ m 2 Fig. 3.
Viability curves for S. cerevisiae irradiated by middle
wavelength UV (290-320 nm): (1) wild type strain; (2) mutant strain rad 3-2. |
|
Viability, %
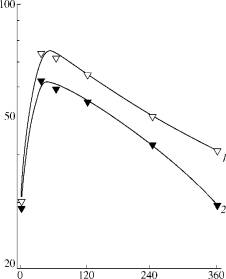
Dose, Jm 2 Fig. 4.
The effect of monochromatic (680 nm) light irradiation dose on photoreduction
of S. cerevisiae cells inactivated by near UV (320-380
nm): (1) wild type strain; (2) mutant strain rad 50-1. Doses of near UV
irradiation: 70 (1) and 30 kJ/m2 (2). |
For this purpose we studied the capacity to FR680 of yeasts C.
cerevisiae, both wild type and its mutants deficient in excision
(rad 3-2) and post-replicative (rad 50-1) DNA reparation. It has been shown
previously that cells of a mutant strains are more sensitive to the near UV
(rad 50-1) and middle wavelength UV (rad-3-2) compared to the wild type strain
(Figs. 2, 3). The photo- reactivation experiments were conducted according to
the following protocol. Both wild type and mutant, rad 50-1 and rad 3-2, cells
were irradiated by fixed doses of near UV or middle wavelength UV, which
decreased their viability to the same level (~30%). After that, they were
irradiated by monochromatic light, 680 nm. The results (Se on Figs. 1,4) show that FR680 in mutant
strains was observed with the same efficiency as in wild type yeasts. This
indicates the lack of excision or postreplicative reparation in the
photorecovery of yeasts inactivated by
UV radiation.
The fact that the effect of a light-sensitive reactive system is not
specific to DNA damage leads to the suggestion that it is possibly involved in
cell recovery from photodamage both in the genome and other cell structures.
Earlier, our studies have shown that under
photodynamic inactivation of yeasts (C. guilliermondii and S. cerevisiae) by high doses of visible light (400-600 nm),
cytoplasm, but not DNA, is the main target [6]. The results presented here, and
which involve the above-described optimal doses at FR680, have shown
that a cell inactivated by visible light can be recovered by monochromatic
irradiation in the wavelength range of 600-730 nm with the highest efficiency
of reactivation at 680 nm. Most important is the fact that the typical curve of
photoreactivation dose in this case has the same shape of curves as an
analogous dose at FR680 for cells inactivated by UV radiation (see
Figs. 1,4). Therefore, the photoreduction of viability of yeast cells
inactivated by visible light involves the same light-sensitive system as that
activated with a lethal dose of UV radiation. Therefore, this inactivation
system functions not only during the induction of various types of DNA damage,
but also in case of photodynamic destruction of plasma membranes.
The similarity in
photoreduction effects for yeast cell viability when they are inactivated both
with UV irradiation and visible light indicates that they are based on an
identical, previously unknown, photo-induced reactivating mechanism, which is
not specific in respect to the nature of lethal damage.
REFERENCES
1. Burchuladze, T.G.
and Fraikin, G.Ya., The Studies of the Mechanism of HADH-sensibilizing
Formation of DNA Nicks at Irradiation by Near UV-light, Mol. Biol.,
1991, vol. 25, no. 4, pp. 955-959.
2. Korogodin, V.I.,
Types of Yeast Cells Inactivation by Ionizing Radiation, Biophysics,
1958, vol. 3, no. 2, pp. 206209.
3. Strakhovskaya,
M.G., Ivanova, E.V., and Fraikin, G.Ya., Stimulating Effect of Serotonin on the
Growth of Yeasts Candida guilliermondii and Bacteria
Streptococcus fac- calis, Microbiology, 1993, vol. 62, no. 1, pp.
46-49.
4. Fraikin, G.Ya.,
Pinyaskina, E.V., Strakhovskaya, M.G., Rubin, A.B., New Photoinduced Defence
System in Candida guilliermondii Cells at Lethal Effect of Middle
Wavelength Ultraviolet Radiation, Dokl. Akad. Nauk, 1995, vol.
343, no. 2, pp. 265-267.
5. Fraikin,
G.Ya., Photoprotection by serotonin, in Biological Effect of Light,
Berlin: New York, 1992, pp. 458-462.
6. Fraikin, G.Ya.,
Strakhovskaya, M.G., and Rubin, A.V., The Role of Membrane Bound Porphyrin-Type
Compound as Endogenous Sensitizer in Photodynamic Damage to Yeast Plasma
Membranes, J. Potochem. Potobiol. B Biol., 1996, vol. 34, no. 1,
pp. 129-135.
7. Peak, J.G., Peak,
M.J., and MacCoss, M., DNA Breakage Caused by 334 nm UV-light is Enhanced by
Naturally Occurring Nucleic Acid Components and Nucleotide Coenzymes,
Photochem. Photobiol., 1984, vol. 39, no. 6, pp. 713-716.
8. Sancar, G.B., DNA
Photolyases: Physical Properties, Action Mechanism and Roles in Dark Repair,
Mutat. Res., 1990, vol. 236, pp. 147-160.