Сизоненко
В.Л., Коваленко Н.И.
Харьковский национальный аграрный университет имени В.В. Докучаева
К плазменной модели распространения нервного импульса
После выхода в свет работ Ходжкина и
Хаксли [1], [2] прошла буквально лавина исследований механизма распространения
нервного импульса в биологически активной мембране (см. [3] – [6] и
цитированную там литературу). В частности, существенное развитие теории этого
явления принадлежит работам Маркина, Чизмаджева и др. авторов, выполненных в
1969–1974 гг. [4, с.373], [5]. Они, по-существу, стали классическими в области
нейробиологии.
В основу теоретических работ было положено
уравнение распространения возбуждения по немиелизированному однородному волокну
мембраны [4, с.373]:
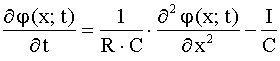 , (1)
, (1)
где ![]() – потенциал мембраны,
отсчитываемый от потенциала покоя,
– потенциал мембраны,
отсчитываемый от потенциала покоя, ![]() – сумма внешнего и
внутреннего сопротивлений,
– сумма внешнего и
внутреннего сопротивлений, ![]() – емкость единицы
длины волокна,
– емкость единицы
длины волокна, ![]() – ионный ток, текущий
через мембрану (отнесенный к единице длины волокна);
– ионный ток, текущий
через мембрану (отнесенный к единице длины волокна); ![]() > 0, если ток
течет наружу.
> 0, если ток
течет наружу.
Если импульс бежит по волокну с постоянной
скоростью ![]() , то уравнение (1), переписанное в терминах переменной
, то уравнение (1), переписанное в терминах переменной ![]() , принимает вид [4]:
, принимает вид [4]:
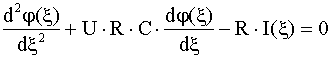 , (2)
, (2)
где выполнены условия: ![]() и
и ![]() . Оказалось, что прямая
. Оказалось, что прямая ![]() = 18,5 мВ пересекает
кривую
= 18,5 мВ пересекает
кривую ![]() в двух точках,
соответствующих большей (
в двух точках,
соответствующих большей (![]() ) и меньшей (
) и меньшей (![]() ) скорости
) скорости ![]() , но одно из решений (
, но одно из решений (![]() ) неустойчиво. Эта теория показала, что скорость
распространения импульса определяется электрическими и геометрическими
параметрами аксона, практические независимо от формы исходного импульса [4].
) неустойчиво. Эта теория показала, что скорость
распространения импульса определяется электрическими и геометрическими
параметрами аксона, практические независимо от формы исходного импульса [4].
Вместе с тем, уравнение (1) является
феноменологическим, выведенным на основании некой электрической схемы мембраны,
и поэтому может не отражать всех тонкостей протекания тока по аксоплазме. В настоящей работе мы развили теорию
протекания токов в цитоплазме аксона и вне го, рассматривая жидкости с обеих
сторон мембраны как электролиты, содержащие плазму ионов разных сортов. В итоге
нами получены уравнения типа (1), (2), но отличающиеся от них в некоторых
деталях.
Будем считать условно, что биологическая
мембрана лежит параллельно оси ![]() в области
в области ![]() и бесконечна вдоль
и бесконечна вдоль ![]() . С наружной стороны мембраны (
. С наружной стороны мембраны (![]() ) присутствует электролит, в котором имеются однозарядные
ионы
) присутствует электролит, в котором имеются однозарядные
ионы ![]() ,
, ![]() и
и ![]() , коэффициенты диффузии которых
, коэффициенты диффузии которых ![]() одинаковы (в действительности они одного порядка):
одинаковы (в действительности они одного порядка):
![]() (
(![]() ) ~
) ~ ![]() (
(![]() ) ~
) ~ ![]() (
(![]() ) ~
) ~ ![]() см2/с, но
для расчетов это не имеет принципиального значения). В цитоплазме аксона (
см2/с, но
для расчетов это не имеет принципиального значения). В цитоплазме аксона (![]() ) находится такой же электролит с ионами
) находится такой же электролит с ионами ![]() ,
, ![]() и
и ![]() , но здесь еще содержится и значительная часть анионов
, но здесь еще содержится и значительная часть анионов ![]() органического происхождения (изэтионовая кислота) [4, с.365],
которые сквозь тело липидного бислоя мембраны проникать не могут. Наличием
двухзарядных ионов
органического происхождения (изэтионовая кислота) [4, с.365],
которые сквозь тело липидного бислоя мембраны проникать не могут. Наличием
двухзарядных ионов ![]() ,
, ![]() и др. пренебрегаем.
и др. пренебрегаем.
Для определения распределения потенциала ![]() электрического поля
электрического поля ![]() в электролите необходимо решить систему уравнений для плотностей
частиц
в электролите необходимо решить систему уравнений для плотностей
частиц ![]() ионы
ионы ![]() ;
; ![]() ;
; ![]() ;
; ![]() ).
).
 , (3)
, (3)
![]() , (4)
, (4)
где
![]()
![]() и
и ![]() – электрический заряд иона
– электрический заряд иона ![]() ,
, ![]() – температура,
– температура, ![]() – диэлектрическая проницаемость среды (
– диэлектрическая проницаемость среды (![]() ≈ 80, как и для воды). Плотности потоков зарядов
≈ 80, как и для воды). Плотности потоков зарядов ![]() определяются
соотношениями
определяются
соотношениями
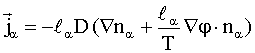 , (5)
, (5)
При этом задаются следующие граничные
условия:
|
|
(6) |
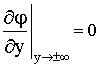
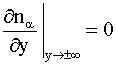
Если учесть, что перенос частиц из-за
диффузии в электролите происходит за характерные времена
![]() ~
~  ~
~ ![]() ~
~ ![]() , (7)
, (7)
которые значительно меньше времени
прохождения нервного импульса (~![]() ), то левые части в (3) малы, и для
), то левые части в (3) малы, и для ![]() справедливы выражения
справедливы выражения
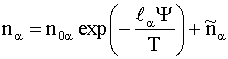 , (8)
, (8)
где
![]() ,
, ![]() при
при ![]() ,
, ![]() при
при ![]() . В области, где
. В области, где ![]() , из (3) находим, что
, из (3) находим, что
 , (9)
, (9)
где
![]() – плотность тока (5)
вдоль оси
– плотность тока (5)
вдоль оси ![]() на поверхности
мембраны
на поверхности
мембраны ![]() ;
; ![]() при
при ![]() ,
, ![]() при
при ![]() . При выводе соотношений (8), (9) предполагалось, что
. При выводе соотношений (8), (9) предполагалось, что ![]() ,
, ![]()
![]()
![]() .
.
Уравнение
(4) с учетом (8) в области ![]() следующее:
следующее:
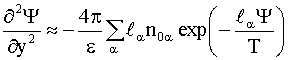 . (10)
. (10)
В случае ![]() решения уравнения
(10) имеют вид:
решения уравнения
(10) имеют вид:
 ;
; 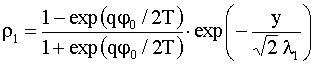 , (11)
, (11)
а при ![]() они заданы
соотношением:
они заданы
соотношением:
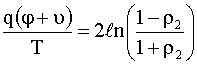 ;
; 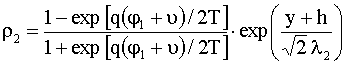 , (12)
, (12)
где
![]() и
и ![]() – значения потенциала
– значения потенциала
![]() на границах мембраны
на границах мембраны ![]() и
и ![]() соответственно:
соответственно:
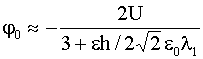 ,
, 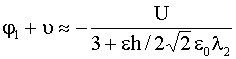 , (13)
, (13)
![]() – радиус Дебая-Гюхкеля в области
– радиус Дебая-Гюхкеля в области ![]() ,
, ![]() – его значение при
– его значение при ![]() ,
, ![]() – диэлектрическая
проницаемость липидного бислоя. При получении соотношений (13) предполагалось,
что
– диэлектрическая
проницаемость липидного бислоя. При получении соотношений (13) предполагалось,
что ![]() , но ими можно пользоваться и в случае
, но ими можно пользоваться и в случае ![]() ~
~ ![]() .
.
На начальной стадии нервного импульса,
когда открываются натриевые каналы, и ионы ![]() начинают входить
внутрь аксона, можно считать, что в наружной среде
начинают входить
внутрь аксона, можно считать, что в наружной среде ![]() ,
, ![]() , а ионы калия и анионы
, а ионы калия и анионы ![]() отсутствуют
отсутствуют ![]()
![]() . Тогда из уравнения (9) с учетом (11) получим, что
. Тогда из уравнения (9) с учетом (11) получим, что ![]() ,
,  ,
,
|
|
(14) |

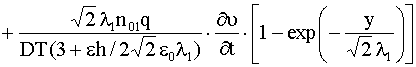
Если выполнено условие
 , (15)
, (15)
то из (14) следует, что в области ![]() плотность
плотность ![]() становится исчезающее
малой
становится исчезающее
малой ![]() , и ток ионов натрия отсутствует.
, и ток ионов натрия отсутствует.
Поскольку ионы хлора (![]() ) не могут проходить через мембрану, то они с уходом ионов
натрия начинают перемещаться в область больших положительных значений
) не могут проходить через мембрану, то они с уходом ионов
натрия начинают перемещаться в область больших положительных значений ![]() . При этом в интервале
. При этом в интервале ![]() из (9) получаем, что
из (9) получаем, что
|
|
(16) |

 .
.В области ![]() плотность
плотность ![]() возрастает с ростом
возрастает с ростом ![]() , достигая при
, достигая при ![]() ~
~ ![]() максимальной величины, которая остается значительно меньшей
максимальной величины, которая остается значительно меньшей ![]() . Здесь же, согласно (4), появляется и маленькое
электрическое поле, которое для наших расчетов не имеет значения. Иными
словами, излишние ионы хлора уходят от поверхности мембраны во внешнюю область,
так что максимум их плотности находится в точке
. Здесь же, согласно (4), появляется и маленькое
электрическое поле, которое для наших расчетов не имеет значения. Иными
словами, излишние ионы хлора уходят от поверхности мембраны во внешнюю область,
так что максимум их плотности находится в точке ![]() ~
~ ![]() .
.
При поступлении ионов натрия во внутреннюю
область аксона уравнения (8), (9) для ![]() здесь неприменимы,
поскольку на какое-то время (порядка
здесь неприменимы,
поскольку на какое-то время (порядка ![]() ~
~ ![]() ) плотность этих ионов может
стать больше исходной равновесной
) плотность этих ионов может
стать больше исходной равновесной ![]() . Однако, при больших временах
. Однако, при больших временах ![]() излишние ионы
излишние ионы ![]() уходят от поверхности
мембраны во внутреннюю часть аксона, так что их плотность
уходят от поверхности
мембраны во внутреннюю часть аксона, так что их плотность ![]() вблизи липидного
бислоя опять становится малой, и на распределение потенциала вдоль
вблизи липидного
бислоя опять становится малой, и на распределение потенциала вдоль ![]() не влияет.
не влияет.
Рассмотрим теперь поведение ионов во
внутренней аксоплазме на второй стадии процесса, когда натриевые каналы
мембраны закрываются, но открываются каналы для ионов ![]() . Тогда
. Тогда ![]() ,
, ![]() ,
, ![]() ,
, ![]() , и для
, и для ![]() становятся
справедливыми соотношения типа (8), (9):
становятся
справедливыми соотношения типа (8), (9):
![]() =
= 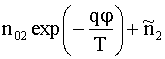 ,
, ![]() , (17)
, (17)
 (18)
(18)
Подставляя в (18) выражение (12) для ![]() , получим аналогично (14), (15) следующие формулы:
, получим аналогично (14), (15) следующие формулы:
 ,
,
|
|
(19) |
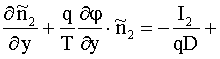
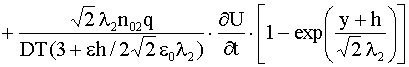 .
.Если выполнено условие
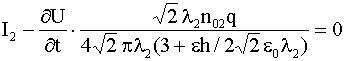 , (20)
, (20)
то из (19) следует, что в области ![]() плотность
плотность ![]() становится исчезающее
малой
становится исчезающее
малой ![]() , и ток ионов калия отсутствует.
, и ток ионов калия отсутствует.
Поскольку анионы ![]() не могут проходить
через мембрану, то они с уходом ионов калия начинают перемещаться в область
больших отрицательных значений
не могут проходить
через мембрану, то они с уходом ионов калия начинают перемещаться в область
больших отрицательных значений ![]() . При этом в интервале
. При этом в интервале ![]() из (9) получаем, что
из (9) получаем, что
|
|
(21) |

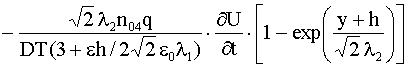 .
.В области ![]() плотность
плотность ![]() возрастает с
уменьшением
возрастает с
уменьшением ![]() , достигая при
, достигая при ![]() ~
~ ![]() максимальной
величины, которая остается значительно меньшей
максимальной
величины, которая остается значительно меньшей ![]() . Здесь же, согласно
(4), возникает и маленькое электрическое поле, которое для наших расчетов не
имеет значения. Иными словами, излишние анионы
. Здесь же, согласно
(4), возникает и маленькое электрическое поле, которое для наших расчетов не
имеет значения. Иными словами, излишние анионы ![]() уходят от поверхности
мембраны внутрь аксона, так что максимум их плотности находится в точке
уходят от поверхности
мембраны внутрь аксона, так что максимум их плотности находится в точке ![]() ~
~ ![]() (
(![]() – момент времени,
когда
– момент времени,
когда ![]() ).
).
При поступлении ионов калия во внешнюю
область аксона уравнения (8), (9) для ![]() здесь неприменимы,
поскольку на какое-то время (порядка
здесь неприменимы,
поскольку на какое-то время (порядка ![]() ) плотность этих ионов может стать больше исходной
равновесной
) плотность этих ионов может стать больше исходной
равновесной ![]() . Однако, при больших
временах (
. Однако, при больших
временах (![]() ) излишние ионы
) излишние ионы ![]() уходят от поверхности
мембраны во внешнюю часть аксона, так что их плотность
уходят от поверхности
мембраны во внешнюю часть аксона, так что их плотность ![]() вблизи липидного
бислоя опять становится малой, и на распределение потенциала вдоль
вблизи липидного
бислоя опять становится малой, и на распределение потенциала вдоль ![]() не влияет.
не влияет.
![]() Основную ценность для нас представляют соотношения
(15) и (20), поскольку они связывают ток через мембрану с изменением потенциала
Основную ценность для нас представляют соотношения
(15) и (20), поскольку они связывают ток через мембрану с изменением потенциала![]() . Полагая далее, что
. Полагая далее, что
|
I = |
|
(22) |
|
|
![]() ,
, ![]() , (23)
, (23)
приходим к следующей формуле связи тока ![]() с потенциалом
с потенциалом ![]() :
:
|
|
при |
(24) |
|
|
при |
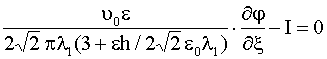 ,
,
Сравним теперь формулы (24) с (2). Видно,
что у нас первые слагаемое в (2) отсутствует, а для емкости ![]() справедлива оценка
справедлива оценка
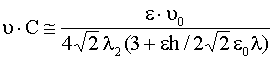 . (25)
. (25)
Прежде, чем сравнивать наши результаты с
экспериментами, отметим, что развитая здесь теория пригодна только для
немиелизированных аксонов, так как мы не предполагали наличие перехватов Ранвье
во внешней области аксона. Как известно, скорости распространения импульсов в
таких системах лежат в пределах ![]() = 0,7 – 2,3 м/с, в то
время как в миелизированных волокнах
они равны:
= 0,7 – 2,3 м/с, в то
время как в миелизированных волокнах
они равны: ![]() = 10–100 м/с [4, c.361]. Беря теперь значения:
= 10–100 м/с [4, c.361]. Беря теперь значения: ![]() ~ 1019 см-3,
~ 1019 см-3, ![]() 30 нм,
30 нм, ![]() = 50 нм,
= 50 нм, ![]() 80,
80, ![]() 4,
4, ![]() = 20 м/с,
= 20 м/с, ![]() = 1,5 м/с, получим из
(25), что
= 1,5 м/с, получим из
(25), что
![]() ~
~ ![]() ед.
ед. ![]() = 0,2 мкΦ/см. (26)
= 0,2 мкΦ/см. (26)
Эта оценка хорошо соответствует значению ![]() = 0,157 мкΦ/см,
найденному для аксонов [4, c.373].
= 0,157 мкΦ/см,
найденному для аксонов [4, c.373].
Нашей моделью можно пользоваться, однако,
и в случае миелизированных волокон, если под током ![]() подразумевать
значение тока, усредненное по некоторой длине, содержащей несколько перехватов
Ранвье. При этом нужно учитывать, что в немиелизированном аксоне через 1 мкм2
поверхности проходит около 20 000 ионов
подразумевать
значение тока, усредненное по некоторой длине, содержащей несколько перехватов
Ранвье. При этом нужно учитывать, что в немиелизированном аксоне через 1 мкм2
поверхности проходит около 20 000 ионов ![]() , так что плотность ионного тока в перехвате примерно в
десять раз больше, чем в немиелизированных гигантских аксонах [4, c.368]. Поэтому для применимости нашей модели ток
, так что плотность ионного тока в перехвате примерно в
десять раз больше, чем в немиелизированных гигантских аксонах [4, c.368]. Поэтому для применимости нашей модели ток ![]() в (24) должен быть
уменьшен приблизительно в десять раз, что эквивалентно увеличению значение
в (24) должен быть
уменьшен приблизительно в десять раз, что эквивалентно увеличению значение ![]() тоже в десять раз,
т.е. до предела
тоже в десять раз,
т.е. до предела ![]() 10
10![]() 15 м/с. В этом случае опять получаем для
15 м/с. В этом случае опять получаем для ![]() оценку (26).
оценку (26).
Уменьшение коэффициента при ![]() во второй формуле
(24) почти в два раза, казалось бы, должно увеличивать крутизну фронта
потенциала
во второй формуле
(24) почти в два раза, казалось бы, должно увеличивать крутизну фронта
потенциала ![]() . В действительности же этого не происходит по той причине,
что ток ионов калия возрастает намного медленнее, чем входной ток ионов натрия.
. В действительности же этого не происходит по той причине,
что ток ионов калия возрастает намного медленнее, чем входной ток ионов натрия.
Развитая нами теория, основанная на
решении точных уравнений диффузии ионов в электрометах, показывает, что первое
слагаемое в (2) привнесено туда искусственно. Однако, без него далеко идущие
выводы о поведении нервных импульсов в аксонах [4], [7], оказываются
недоказанными.
Литература:
1.
Hodgkin A.L., Huxley A.F. // I. Physiol. 1952. V. 117. P. 500–544.
2.
Ходжкин А. Нервный
импульс: Пер. с англ. М.: Мир, 1965.
3.
Шеперд Г. Нейробиология.
Т. 1: Пер. с англ. М.: Мир, 1987. – 454 с.
4.
Волькенштейн М.В.
Биофизика. М.: Наука, 1988. – 592 с.
5.
Ходоров Б.И. Общая
биология возбудимых мембран. М.: Наука, 1975.
6.
Тасаки Н. Нервное
возбуждение: Пер. с англ. М.: Мир, 1971.
7.
Маркин В.С., Пастушенко
В.Ф., Чизмаджев Ю.А. Теория возбудимых сред. М.: Наука, 1981